|
Pictures by Chad Thomas, Texas State University-San Marcos |
||
|
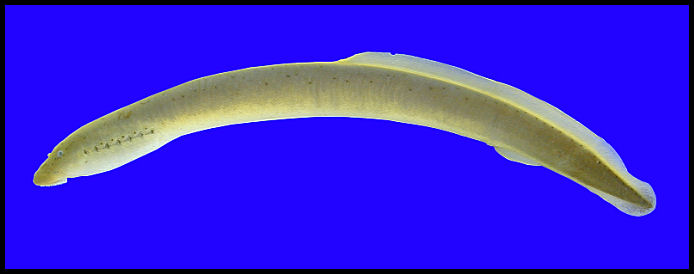
southern brook lamprey
Type Locality Stream 0.8 km (0.5 mi) south of Dry Prong, Grant Par., LA (Hubbs and Trautman 1937)
Etymology/Derivation of Scientific Name Ichthyomyzon, Greek, “fish sucker”; gagei, named for S.H. Gage, one of the foremost students of Lampreys (Pflieger 1997).
Synonymy Ichthymyzon gagei Hubbs and Trautman 1937:80; Cook 1959:45.
Characters Maximum size: 166 mm (6.5 in) TL (Knapp 1951).
Coloration: It is gray to brown dorsally, becoming lighter tan or cream colored along the sides. The tail and head are generally darker than midbody regions. Lateral line is usually (but not always) conspicuous with dark spots or as lighter rings surrounded by a dark rim (Ross 2001).
Counts: Trunk myomeres 50-52 (49-50), supraoral cusps 3 (2-5), and infraoral cusps number 5-12 (Ross 2001).
Body shape: Eel-like.
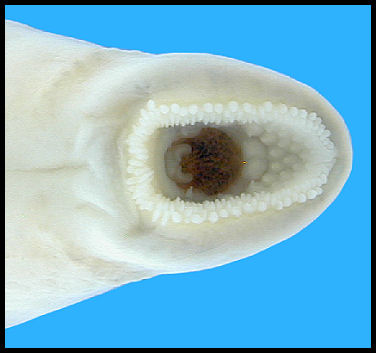
Mouth position: Subterminal (Goldstein and Simon 1999). Teeth degenerate (Dendy and Scott 1953).
Morphology: Mouth disc small; length of disc contained 17.2-26.3 times in total length; cusps in posterior field degenerate (Hubbs et al. 1991); eye well-developed in adults (Ross 2001). Intestine not developed (Hubbs et al. 1991).
Distribution (Native and Introduced) U.S. distribution: Ranges along Gulf of Mexico drainages from western Florida to eastern Texas, north through eastern Oklahoma to Missouri, and east to Tennessee (Hubbs et al. 1991). Disjunct population reported to inhabit the upper Mississippi River in Wisconsin and Minnesota (Cochran 1987).
Texas distribution: Occurs in Red, Sabine, San Jacinto, and Neches basins (Hubbs et al. 1991).
[Additional literature noting collection of this species from Texas locations includes, but is not limited to the following: Hubbs (1957); Big Sandy Creek (Evans and Noble 1979).]
Abundance/Conservation status (Federal, State, Non-governmental organizations) Populations in the southern United States are currently secure (Warren et al. 2000).
Habitat Associations Macrohabitat: Streams (Rhode and Lanteigne-Courchene 1980; Moshin and Gallaway 1977).
Mesohabitat: Restricted to freshwater (Hardisty and Potter 1971). Adults occur in fairly swift water (30-50 cm/s) over a coarse-sand substrate, although adults may also occur in the slow current area more typical of ammocoetes (Beamish 1982; Beamish and Thomas 1984). Ammocoetes are found in sand substrate of small streams and occasionally in accumulations of silt and decaying leaves adjacent to sand bars (Dendy and Scott 1953; Carlander 1969).
Biology Spawning season: In Mill Creek drainage, Texas, most of reproduction occurred in the spring; however sampling data suggests that some spawning may occur over a longer period (Moshin and Gallaway 1977). In Choclafaula Creek, Alabama, adults spawned between mid-April and early May (William and Beamish 1982).
Spawning habitat: Gravel or sand; Dendy and Scott (1953) found 3-4 nests on the upstream end of a riffle where the bottom material was gravel and sand, in water 51-76 mm (2-3 in) deep; largest pebbles were about 51 mm (2 in) in diameter; nests consisted of round depressions 152.4-203.2 mm (6-8 in) wide and approximately 51 mm (2 in) deep.
Reproductive strategy: Dendy and Scott (1953) observed spawning on Choclafaula Creek (Alabama River System): the number of lampreys participating in nest building and spawning activities ranged from 5-20 or more per nest. Lampreys carrying pebbles with their sucking discs swam rapidly in an upstream or lateral direction removing the pebbles from the nest. On one occasion two individuals attached themselves to one stone which was too large for a single lamprey to move and thus succeeded in removing it. This behavior may have been accidental cooperation. Some lampreys moved from one nest to another, thus, giving the nesting an appearance of a community project. Dendy and Scott (1953) observed and described reproductive behavior in aquarium conditions.
Fecundity: In an Alabama population, number of oocytes ranged from 820-2,485; maximum gonadosomic ratio in males was 17.1% and 26.3% in females (William and Beamish 1982). Egg counts prior to spawning for 10 females ranged from 1,000-3,264 with an average of 1,787 (Dendy and Scott 1953). Hatching occurs 2-3 weeks after spawning (Beamish 1993). Beamish et al. (1994) found that fecundity was subject to environmental influence. Age at maturation: Ammocoetes require at least 3 years to complete development (Rhode and Lanteigne-Courchene 1980). Ammocoetes as small as 80 mm (3 in) TL may have well-developed gonads (Boschung and Mayden 2004).
Migration: During transformation, young lampreys migrate to clear, moderately flowing waters over sand and relatively clean gravel where transformation is completed (Dendy and Scott 1953; Boschung and Mayden 2004).
Longevity: Adults die within 2-26 days after spawning (Dendy and Scott 1953).
Growth and Population Structure: Recently hatched ammocoetes 11-14 mm (.43-.55 in) TL (Moshin and Gallaway 1977). Ammocoetes require at least three years to complete development (Rohde and Lanteigne-Courchene 1980). In an Alabama study, length-frequency curves for larvae indicated a larval life of 3 ¼- 4 ¼ years. Growth rate was constant throughout the year for each age group. Sex ration of larval I. gagei (male to female) was 1:1.02. Metamorphosis initiated in early September, completed between early February and mid-March (William and Beamish 1982). Boschung and Mayden (2004) report collection of adults as small as 82 mm (3.2 in) TL. Beamish and Thomas (1984) reported large ammocoetes beginning to metamorphosis to adult stage in late August or early September, duration of metamorphosis approximately 6 months; minimum sizes at metamorphosis at least 100 mm (4 in) for males and 105 (4.1 in) mm for females. Females tend to be longer then males, with females averaging 117-127 mm (4.6-5.0 in) TL compared to 106-116 mm (4.2-4.6 in) TL for males (Dendy and Scott 1953; Moshin and Gallaway 1977; Beamish 1982). In Mill Creek drainage, Texas, ammocoetes were abundant during all seasons, while adults were relatively abundant only during winter and spring periods (Moshin and Gallaway 1977). Sexes of ammocoetes not distinct until after about 50-90 mm (2.0-3.5 in), when the larvae are 12-17 months of age (Ross 2001); rapid development of eggs in almost all larvae of this size range, but eggs developing in “males” were then reabsorbed; occurrence of oocytes in testes of large male not uncommon (Beamish and Thomas 1983). Environmental factors including average growth rate of individuals in a population, larval density, pH, average stream temperature largely control sex determination (Beamish 1993).
Food habits: Herbivore/detritivore; filter feeder; filterer (trophic mode; Goldstein and Simon 1999). Ammocoetes feed on minute organic particles for several years prior to metamorphosing into adults (Carlander 1969). Southern brook lampreys do not feed following the larval interval, but relies on stored energy to sustain their metabolic requirements, including gonadal maturation, through to the completion of spawning after which they die (Beamish and Legrow 1983). Main food items include plankton, particularly diatoms and organic detritus (Moshin and Gallaway 1977). In Mill Creek drainage, Texas, condition of ammocoetes markedly increased from winter to spring, which coincided with an increase in the amount of food ingested, the organic content of the material, and with presumed changes associated with metamorphosis into adults (Moshin and Gallaway 1977).
Phylogeny and morphologically similar fishes Ichthyomyzon gagei is thought to be derived from the parasitic chestnut lamprey (I. castaneus) to which it is most similar, and from which it differs as an adult by the oral disk being narrower (rather than wider) than head with width and by the degenerate digestive tract. The ammocoetes are very difficult to separate, but I. gagei tends to have less pigmentation below the eye and in front of the gill region and usually has well pigmented lateral line organs. Because of rapid sexual development at the end of the larval period, late stage ammocoetes (greater than 100mm, 4 in) often have well developed gonads. The presence of developed gonads in the ammocoetes stage is the best means of distinguishing I. gagei from the I. castaneus (Ross 2001).
Host Records No information at this time.
Commercial or Environmental Importance No information at this time.
References Beamish, F.W.H. 1982. Biology of the southern brook lamprey, Ichthyomyzon gagei. Env. Bio. Fish. 7(4): 305-320. Beamish, F.W.H. 1983. Potential and actual fecundity of the “paired” lampreys, Ichthyomyzon gagei and I. castaneus. Copeia 1983(2):367-374. Beamish, F.W.H. 1993. Environmental sex determination in southern brook lamprey, Ichthyomyzon gagei. Can. J. Fish. Aquat. Sci. 50:1299-1307. Beamish, F.W.H., and M. Legrow. 1983. Bioenergetics of the southern brook lamprey, Ichthyomyzon gagei. Journal of Animal Ecology 52:575-590. Beamish, F.W.H. and E.J. Thomas. 1984. Metamorphosis of the southern brook lamprey, Ichthyomyzon castaneus. Copeia 1984(2): 502-515. Beamish, F.W.H., P.K. Ott, S.L. Roe. 1994. Interpopulational variation in fecundity and egg size in southern brook lamprey, Ichthyomyzon gagei. Copeia 1994(3):718-725. Boschung, H.T., Jr., and R.L. Mayden. 2004. Fishes of Alabama. Smithsonian Books, Washington. 736 pp. Carlander, K.D. 1969. Handbook of Freshwater fishery biology. Vol.1. The Iowa State Univ. Press, Ames. Cochran, P.A. 1987. The southern brook lamprey (Ichthyomyzon gagei) in the St. Croix River drainage of Wisconsin and Minnesota. Copeia 1987(2):443-446. Cook, F.A. 1959. Freshwater fishes in Mississippi. Mississippi Game and Fish Commission, Jackson. Dendy, J. S. and D.C. Scott. 1953. Distribution, Life History, and Morphological Variations of the Southern Brook Lamprey, Ichthyomyzon gagei. Copeia 1953(3):152-162. Evans, J.W., and R.L. Noble. 1979. The longitudinal distribution of fishes in an east Texas stream. American Midland Naturalist 101(2):333-343. Goldstein, R.M., and T.P. Simon. 1999. Toward a united definition of guild structure for feeding ecology of North American freshwater fishes. pp. 123-202 in T.P. Simon, editor. Assessing the sustainability and biological integrity of water resources using fish communities. CRC Press, Boca Raton, Florida. Hardisty, W.M. and I.C. Potter. 1971. The behavior, ecology and growth of larval lampreys, pp 85-125. In biology of lampreys. Vol. 1. M. W. Hardisty and I.C. Potter, eds. Academic Press, New York, NY. Hubbs, C. 1957. Distributional patterns of Texas fresh-water fishes. The Southwestern Naturalist 2(2/3):89-104. Hubbs, C.L. and M. B. Trautman. 1937. A revision of the lamprey genus Ichthyomyzon. Misc. Publ. Mus. Zool. Univ. Mich. 35:1-109. Hubbs, Carl L., R.J. Edwards and G.P. Garret. 1991. An annotated checklist of freshwater fishes of Texas, with key to identification of species. Texas Journal of Science, Supplement 43(4):1-56. Knapp, F. T. 1951. Additional Reports of Lampreys from Texas. Copeia 1951(1):87. Moshin, A.K.M., and B.J. Gallaway. 1977. Seasonal abundance, distribution, food habitats and condition of the southern brook lamprey, Ichthyomyzon gagei Hubbs and Trautman, in an east Texas watershed. The Southwestern Naturalist 22(1):107-114. Pflieger, W.L. 1997. The Fishes of Missouri. Revised Edition. Missouri Department of Conservation. Jefferson City. 372 pp. Rohde, F.C. and J. Lanteigne-Courchene 1980. Ichthyomyzon gagei (Hubbs and Trautman), Southern Brook Lamprey.pp.18 in D.S. Lee et al. Atlas of North American Freshwater Fishes. N.C. State Mus. Nat. Hist., Raleigh, i-r+854 pp. Ross, Stephen T. 2001. The Inland Fishes of Mississippi. University Press of Mississippi, Jackson. 624 pp. Warren, L. W., Jr., B. M. Burr, S. J. Walsh, H. L. Bart, Jr., R. C. Cashner, D. A. Etnier, B. J. Freeman, B. R. Kuhajda, R. L. Mayden, H. W. Robison, S. T. Ross, and W. C. Starnes. 2000. Diversity, Distribution, and Conservation status of the native freshwater fishes of the southern United States. Fisheries 25(10):7-29. William, F., and H. Beamish. 1982. Biology of the southern brook lamprey, Ichthyomyzon gagei. Environmental Biology of Fishes 7(4):302-320.
|
||
|
|
||